Research & Teaching
Current research projects:
- Investigating the roles of error-prone DNA polymerases in double-strand break repair.
- Determining the genetic components and molecular mechanisms of alternative end-joining repair pathways.
- Using inducible nuclease systems to determine how DNA sequences adjacent to a double-strand break regulate preferred repair outcomes.
- Identifying the hierarchy of proteins and pathways that are used during tolerance of DNA damage.
- Elucidating the relationship between structure-forming DNA sequences, the proteins that remove them, and how they can induce genome instability.
- Identifying interventions that can extend the healthspan of Drosophila and other eukaryotic organisms.
Watch three undergraduate students describe why they do research and talk about their projects
Focus on DNA damage and repair

In order to accurately replicate and pass on their genetic material, cells must repair DNA damage as it arises. Two of the most dangerous types of DNA damage are double-strand breaks and interstrand crosslinks. Failure to repair these lesions can result in cell death by apoptosis, while inaccurate repair can be mutagenic. Many human diseases, including Fanconi Anemia, Bloom Syndrome, and other cancer-prone disorders, are caused by defects in repair mechanisms that deal with DNA breaks and crosslinks.
The McVey lab characterizes processes involved in repair of DNA breaks and crosslinks in the model metazoan, Drosophila melanogaster. Our long-term goal is to determine how various DNA repair pathways are regulated during development and aging in a multicellular eukaryote.
Our research employs a variety of genetic and molecular approaches, including powerful assays in which we can create double-strand breaks at known sites in the genome and recover and molecularly analyze repair events. The phenotypic results of one such assay are shown in Figure 1. Because Drosophila is a such a superb genetic system, we can use a wide array of tools to alter genes and gene expression patterns in order to test various DNA repair models.
Tolerating DNA damage
When cells are actively copying their DNA, the normal replicative DNA polymerases often get 'stuck' when they arrive at a damaged region of the template. In order to continue the copying process, they use damage tolerance mechanisms to get past the damage. Options include skipping over the damage and repriming further downstream on the template, using specialized translesion DNA polymerases to copy the damaged template, and reversing the replication fork to give the replication machinery time and space to deal with the damage. We're interested in the ways that cells choose between these options. We have recently identified Rev1 as a protein that is vital to the decision-making process. Ongoing projects in the lab are poised to elucidate how Rev1 interacts with other proteins to regulate damage tolerance.
Homologous recombination repair
Double-strand breaks can be repaired by two main classes of pathways: non-homologous end-joining and homologous recombination. Homologous recombination (HR) utilizes a homologous template for repair and is usually considered to be error-free. However, experiments done using budding yeast suggest that HR can be mutagenic in certain contexts. We have demonstrated that error-prone, translesion polymerases, such as polymerase eta, polymerase zeta, and Rev1, have important roles during HR repair in flies. We are now testing the hypothesis that HR is also mutagenic in metazoans and that translesion DNA polymerases are responsible for at least a portion of this mutagenesis.
Non-homologous end joining
End joining represents a flexible set of mechanisms that can repair double-strand breaks when a homologous template is unavailable. Classical end joining (C-NHEJ) involves the protection, processing, and subsequent ligation of broken ends, and depends on the Ku70/80 heterodimer and the DNA ligase IV/XRCC4 complex. Cells lacking C-NHEJ proteins can utilize one or more alternative end-joining (alt-EJ) mechanisms. Currently, there is much intrest
We have found that Drosophila polymerase theta (pol theta) is involved in a particular form of alt-EJ that we have termed synthesis-dependent microhomology-mediated end joining (see Figure 2). Interestingly, pol theta is also involved in DNA interstrand crosslink repair. We are further characterizing these dual roles of pol theta using biochemical and molecular biology approaches. The relevance of our research to human health is highlighted by recent research showing that pol theta is highly expressed in several different types of human tumors and that overexpression correlates with poor prognosis. Could DNA polymerase theta be a therapeutic target? Many research groups are now pursuing this avenue of research.
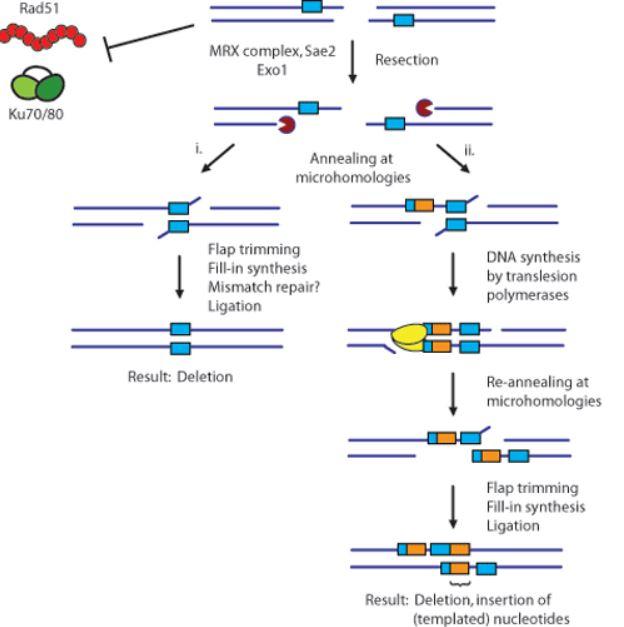
Figure 2: Two potential pathways of alternative end joining. (i) microhomology-mediated end joining, which involves annealing at pre-existing microhomologous sequences. (ii) synthesis-dependent microhomology-mediated end joining, which involves DNA synthesis and production of nascent microhomologies.